Clase #2: Células Eucariotas y Células Procariotas
CELULA EUCARIOTA
Se
conoce como célula eucariota a aquellas en cuyo citoplasma puede
hallarse un núcleo
celular que contiene el material genético (ADN), a diferencia
de las células procariotas, cuyo material genético se halla disperso en el
citoplasma.
La
aparición de este tipo de células se considera un importante paso evolutivo ya
que sentó las bases para la futura complejidad y variedad de la vida
pluricelular, dando origen así a los reinos superiores (animalia, plantae, fungi y protistae).
Los
seres vivos formados por células eucariotas se denominan eucariontes.
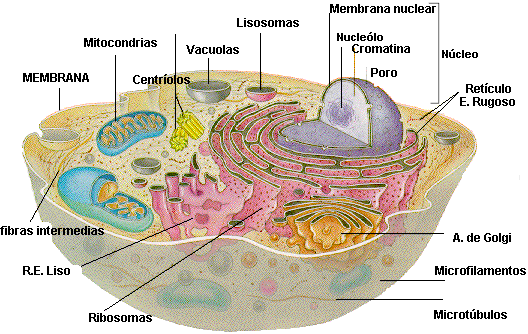
ESTRUCTURA DE CÉLULAS EUCARIOTAS
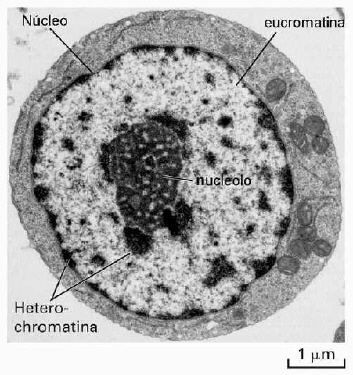
Núcleo.-
El núcleo contiene el genoma de la célula. Está
limitado por una membrana formada por un par de unidades de membrana separadas
por un espacio de grosor variable. La membrana interna por lo común es un saco
simple, pero la membrana más externa se presenta en varios sitios como una
continuación del retículo endoplásmico. La membrana nuclear muestra
permeabilidad selectiva por la presencia de poros, que consisten en varias
proteínas complejas cuya función es importar sustancias y extraer sustancias
del núcleo. Los cromosomas de las células eucariotas contienen macromoléculas
de DNA lineal expuestas en una doble hélice. Sólo son visibles con la
microscopia de luz cuando la célula se encuentra en división y el DNA se
encuentra en una forma muy condensada; en otros momentos los cromosomas no se encuentran
condensados y tienen el aspecto que se muestra en la fi gura 2-2. Las
macromoléculas de DNA de las células eucariotas se asocian con proteínas
básicas denominadas histonas que se unen al DNA por medio de interacciones iónicas.
Una estructura a menudo visible en el núcleo es el nucléolo, un área rica en RNA
que es el sitio de síntesis del RNA ribosómico (fi g. 2-2). Las proteínas
ribosómicas sintetizadas en el citoplasma se transportan hacia el nucléolo y se
combinan con RNA ribosómico para formar subunidades grandes y pequeñas de ribosoma
eucariota. Más tarde éstas son llevadas al citoplasma donde se asocian para
formar ribosomas intactos que participan en la síntesis de proteínas.
ESTRUCTURAS
CITOPLÁSMICAS
El
citoplasma de las células eucariotas se caracteriza por la presencia de un
retículo endoplásmico, vacuolas, plástidos que se reproducen por sí mismos y un
citoesqueleto complejo, compuesto por microtúbulos, microfi lamentos y fi lamentos
intermedios.
El retículo endoplásmico (ER, endoplasmic
reticulum) es una red de conductos limitados por membranas que tienen
continuidad con la membrana del núcleo. Se identifican dos tipos de retículo
endoplásmico: rugoso, al cual se unen ribosomas 80S y liso, sin dichos
ribosomas (fig. 2-2). El retículo endoplásmico rugoso es el principal productor
de glucoproteínas y también produce nuevo material de membrana que se
transporta a través de la célula; el retículo endoplásmico liso participa en la
síntesis de lípidos y en algunos aspectos del metabolismo de los carbohidratos.
El aparato de Golgi consiste en un conjunto de membranas que funcionan en
combinación con el retículo endoplásmico para modifi car y organizar productos
químicos del retículo endoplásmico que más tarde serán secretados y aquellos que
participan en la producción de otras estructuras de la membrana celular.
Los
plástidos incluyen mitocondrias y cloroplastos. Varias pruebas sugieren que las
mitocondrias y cloroplastos son descendientes de microorganismos procariotas
antiguos y que se originaron del englobamiento de células procariotas por células
de mayor tamaño (endosimbiosis). El retículo endoplásmico (ER, endoplasmic
reticulum) es una red de conductos limitados por membranas que tienen
continuidad con la membrana del núcleo. Se identifi can dos tipos de retículo
endoplásmico: rugoso, al cual se unen ribosomas 80S y liso, sin dichos
ribosomas.

CELULA PROCARIOTA
ESTRUCTURA DE LAS CÉLULAS PROCARIOTAS
La
célula procariota es más simple que la eucariota al nivel de la Energía, con
una excepción: la envoltura celular es más compleja.
Nucleoide
Las
células procariotas no tienen un núcleo verdadero; almacenan su DNA en una
estructura conocida como nucleoide. El nucleoide puede observarse en la
microscopia de luz en material teñido (fi g. 2-4). Es positivo para el
colorante de Feulgen, lo que indica la presencia de DNA. El DNA de carga
negativa es neutralizado, al menos en parte, por poliaminas pequeñas y por
iones de magnesio, pero las proteínas similares a histonas existen en bacterias
y tal vez desempeñan una función similar a la de las histonas en la cromatina
de las células eucariotas.
Estructuras citoplásmicas
Las
células procariotas carecen de plástidos autónomos, como las mitocondrias y
cloroplastos; enzimas de transporte de
electrones se localizan en la membrana citoplásmica. Los pigmentos fotosintéticos
(carotenoides, bacterioclorofi la) de bacterias fotosintéticas se encuentran contenidos
en sistemas de membranas intracitoplásmicas de varias morfologías. Las vesículas
de membrana (cromatóforos) son tipos de membrana observadas a menudo. Algunas bacterias
fotosintéticas tienen estructuras especializadas rodeadas por membrana denominadas
clorosomas.
Envoltura celular
Las
células procariotas están rodeadas por una envoltura compleja en capas que difi
ere en composición entre los principales grupos. Tales estructuras protegen al
microorganismo de entornos ambientales hostiles, como osmolaridad extrema,
químicos nocivos e incluso antibióticos.
Membrana celular
Estructura
Las
membranas celulares de las arqueobacterias difieren delas de las bacterias. Las
membranas celulares de algunas arqueobacterias contienen un lípido singular,
los isoprenoides en lugar de ácidos grasos, unidos al glicerol por un enlace
éter en lugar de un enlace éster. Algunos de estos lípidos no contienen grupos
fosfato y por tanto no son fosfolípidos.
Función
Las
principales funciones de la membrana citoplásmica son: 1) permeabilidad
selectiva y transporte de solutos; 2) transporte de electrones y fosforilación
oxidativa en especies aerobias; ex-concreción de exoenzimas hidrolíticas; 4)
transporte de enzimas y moléculas que participan en la biosíntesis de DNA,
polímeros de la pared celular y lípidos de la membrana, y 5) portar receptores y
otras proteínas quimiotácticas y otros sistemas sensoriales de transducción.
Pared celular
La
presión osmótica interna de la mayor parte de las bacterias varía de 5 a 20 atm
como consecuencia de la concentración de solutos por medio del transporte
activo. En la mayor parte de los entornos, esta presión sería sufi ciente para
hacer estallar la célula si no se contara con la presencia de una pared celular
con fuerza tensil elevada (fi g. 2-13). La pared celular bacteriana debe su
resistencia a una capa compuesta de diversas sustancias conocidas como mureína,
mucopéptidos o peptidoglucanos (todos son sinónimos). La estructura de los
peptidoglucanos se revisa adelante.
Capa de peptidoglucanos
El
peptidoglucano es un polímero complejo que consiste, con fines de descripción,
de tres partes: una

Endosporas
Miembros
de varios géneros bacterianos son capaces de formar endosporas (fi g. 2-27).
Las dos más comunes son bacilos grampositivos: los anaerobios obligados del
género Bacillus y los anaerobios obligados del género Clostridium. Otras
bacterias que se sabe forman endosporas son Th ermoactinomyces, Sporolactobacillus,
Sporosarcina, Sporotomaculum, Sporomusa y Sporohalobacter. Dichos
microorganismos sufren un ciclo de diferenciación en respuesta a condiciones
ambientales: el proceso, denominado esporulación, es desencadenado por el casi agotamiento
de varios nutrientes (carbono, nitrógeno o fósforo).
Esporulación
El
proceso de esporulación inicia cuando las condiciones nutricionales se tornan
poco favorables, hay casi agotamiento de las fuentes de nitrógeno o de carbono
(o ambas), lo que constituye los factores más significativos. La esporulación
ocurre masivamente en cultivos que han terminado su crecimiento exponencial
como consecuencia del casi agotamiento de los nutrientes. La esporulación
implica la producción de muchas estructuras, enzimas y metabolitos nuevos junto
con la desaparición de varios componentes de la célula vegetativa.




Comentarios
Publicar un comentario